The first extensive examination of neuronal identity was performed by Nicholls and Baylor (1968) on mechanosensory neurons which are sensory neurons that respond to cutaneous stimuli. Mechanosensory neurons were well suited for this study because their somata lie within the segmental ganglion and their axons extend out the nerve roots where they terminate directly in the tegument. No peripheral synapses are involved in processing cutaneous stimuli.
How does one establish the structure - function relationship of neurons in a segmental ganglion? Nicholls and Baylor (1968) addressed this question using a preparation consisting of a flap of body wall that was several segments in length and connected to a segmental ganglion by the nerve roots (Fig. 4.2a). Using this preparation, Nicholls and Baylor (1968) found that each segmental ganglion contains three classes of mechanosensory neurons that respond specifically, and selectively, to various degrees of cutaneous stimulation: touch (T) cells (three pairs per ganglion) respond to light touch of the body wall; pressure (P) cells (two pairs per ganglion) respond following marked deformation of the body wall and nociceptive (N) cells (two pairs per ganglion) respond to noxious stimuli (Figs. 4.2B - 4.2e). The size, shape, and position of the mechanosensory neurons within each segmental ganglion were remarkably stereotyped, as were the size, shape and position of their receptive fields on each body wall segment. Similarly, the physiological properties of each class of mechanosensory neurons were distinct from one another but consistent from one segmental ganglion to the next. Thus each class of mechanosensory neurons had a unique structure - function relationship. More importantly, the results of Nicholls and Baylor (1968) suggested that all 200 pairs of segmental neurons could be identified and their functional role in leech behavior discerned. One of the first behaviors systematically studied at the level of identified neurons was leech swimming.
Two early studies of leech swimming movements provided the ground work for further investigations into the neural basis of swimming. First,ll (1905) showed that disconnecting the head and tail ganglia from the segmental nerve cord did not abolish leech swimming. Hence swimming movements are generated within segmental ganglia. Second, Gray et al. (1938) demonstrated that the neural activity responsible for coordinating swimming occurred via the intersegmental connectives and not through the body wall. A result of these two studies was the development of a semi-intact preparation, where several mid-body segmental ganglia were exposed for intracellular and extracellular recording, while leaving more anterior and posterior body wall regions intact (Kristan et al., 1974). Using semi-intact preparations, and later using isolated nerve cord preparations, the role of specific neurons in the segmental ganglia and subesophageal ganglion, with respect to swimming, was determined (Kristan et al., 1974 and Kristan and Calabrese, 1976). Experiments were designed to identify how peripheral sensory information propagated, neuron-to-neuron, through the nervous system to generate the antiphasic contractions of the dorsal and ventral longitudinal muscles of a swimming leech.
Body wall movements during swimming are controlled by four groups of segmental motor neurons: the ventral excitors (VEs) that excite the ventral longitudinal muscles; ventral inhibitors (VIs) that inhibit the ventral longitudinal muscles and the VEs; dorsal excitors (DEs) that excite the dorsal longitudinal muscles; and dorsal inhibitor (DIs) that inhibit the dorsal longitudinal muscles and the DEs (Ort et al., 1974). The membrane potential of each motor neuron oscillates and fire bursts of action potentials during their depolarized phase (Fig. 4.3b).
At the center of the swim-generating network is an ensemble of neurons that comprise the central pattern generator or oscillator which provides the appropriate intra- and intersegmental phasic input to segmental motor neurons that produce the undulatory body wall movements characteristic of swimming leeches. Three physiological criteria were used to determine if a given neuron was a member of the swim oscillator. First, the membrane potential of the neuron had to oscillate in phase with the swim motor pattern. Second, the neuron had to be synaptically connected to other members of the oscillator and lastly, perturbations of membrane potential oscillations in the putative oscillator neuron had to cause a transient phase-shift of the swim motor pattern (Friesen et al., 1978 and Poon et al., 1978). To date, six paired (cells 115, 33, 28, 27, 123, and 60) and one unpaired interneuron (cell 208), and fours pairs of motor neurons (cells 1, 2, 102 and 119) meet these criteria (Friesen, 1989; Fig. 4.4a).
With the identification of a central swim oscillatory network and its output connections to motor neurons, attention turned to understanding how the swim oscillatory network was 'turned-on'. Weeks and Kristan (1978) identified an unpaired intersegmental interneuron, cell 204, located in segmental ganglia 10 - 16 that had 'command-like' properties for the initiation of swimming. In isolated cords of segmental ganglia, depolarization of a single cell 204 such that the cell fires at an average frequency of 20 - 50 Hz, initiated swimming with swimming activity generally lasting as long as a suprathreshold firing frequency was maintained in cell 204 (Fig. 4.4b). Because of this latter property cells 204 were classified as swim-gating interneurons (Friesen, 1989). Cells 204 activate the swim oscillator via direct connections to three oscillator neurons, cells 28, 115 and 208 (Nusbaum et al., 1987). What was still lacking was an understanding of how the swim-gating interneurons were themselves driven. Cutaneous stimuli which elicited swimming only indirectly excited cells 204. A neuron or groups of neurons had to be interposed between the mechanosensory neurons and the swim-gating interneurons.
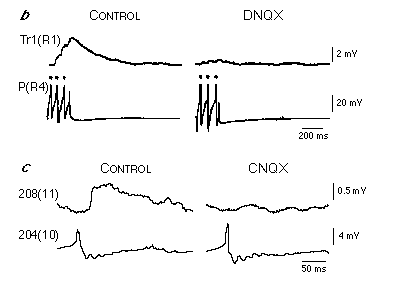
The identification of cells Tr1 provided, in part, the missing link between the mechanosensory input and the activation of swim-gating interneurons. Cells Tr1 are paired interneurons that have their somata in the subesophageal ganglion and extend their axons the length of the ventral nerve cord (Brodfuehrer and Friesen, 1986a,b). They receive direct excitatory input from P and N cells located in the subesophageal ganglion and in the first segmental ganglion, and provide output directly onto all segmental swim-gating interneurons (Brodfuehrer and Friesen, 1986b). Furthermore, brief (approximately 1 s), high frequency (30 to 50 Hz) stimulation of cell Tr1 can elicit a swim episode (Fig. 4.4c), with the length of elicited swim episode being independent of cell Tr1 stimulation intensity. These physiological properties along with the fact that cells Tr1 are silent during swimming led to their classification as swim trigger neurons.
With the identification of cells Tr1 and their input - output connections, the goal of understanding the neuronal basis of leech swimming has been accomplished at one level. That is, at the level of knowing how specific mechanosensory inputs propagate through the leech CNS to activate the swim oscillator and produce swimming movements. Although significant this connectionist model of the leech swim-generating network does not adequately explain the neuronal mechanism governing swimming nor does it explain the behavioral variability observed in the ability of a given input to initiate swimming. A complete understanding of the leech swim-generating network requires a thorough characterization of the biophysical (ionic currents and synaptic transfer functions) and the biochemical (neurotransmitter and receptor phenotypes) properties that govern the synaptic interactions between neurons, and must incorporate behavioral variability into the model. In the next two sections we will discuss recent observations that extend our connectionist understanding of leech swimming to include the roles that the neurotransmitter glutamate and the 'internal state' of the nervous system play in the initiation of swimming.
L-Glutamate and non-NMDA receptors also appear to mediate the synaptic connections from P cells to cell Tr1, and from cell 204 to swim oscillator neurons - cells 208, 28 and 115 (Thorogood and Brodfuehrer, 1995 and Thorogood et al., 1996; Figs. 5B and C). However, the time course of these synaptic events are different than that between cell Tr1 and cell 204. Activation of neither cell Tr1 nor oscillator neurons greatly outlasts the stimulus duration in cell P or cell 204, respectively. This suggests that there exists a functional distribution of glutamate receptors in leech neurons (Dierkes et al., 1996). It is also interesting that L-glutamate plays a role in controlling the initiation of swimming only with respect to the input pathway to the swim oscillator (see Fig. 4.3a). On the output side, the synaptic connections from oscillator neuron cell 208 to DE-3, a dorsal excitor motor neuron, and from DE-3 to the dorsal longitudinal muscles are not blocked by the same non-NMDA receptor antagonists that block glutamatergic connections on the input side (Thorogood et al., 1996).
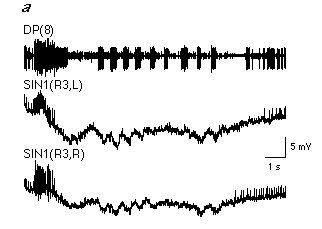
Recent attempts to correlate the activity of individual neurons with the likelihood that a given stimulus will initiate swimming has led to the hypothesis that the control of swimming involves two parallel systems originating in the head ganglion that have opposite effects on the segmental swim-generating network: a swim-activating system that excites the segmental swim-generating network and a swim-inactivating system that inhibits or suppresses it (Brodfuehrer and Burns, 1995). In order for a given stimulus to initiate swimming the swim-activating system must be "turned on" and the inactivating system "turned off". Evidence supporting this dual control mechanism is based primarily on the activity patterns observed in cells 204 and cell SIN1, an identified interneuron in the subesophageal ganglion (Brodfuehrer and Burns, 1995).
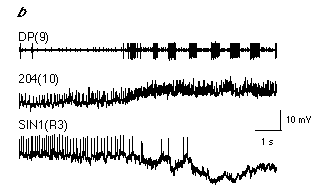
In quiescent preparations (i.e. when swimming is not occurring) cells SIN1 are normally tonically active. When swimming occurs, spiking activity in cells SIN1 ceases and their membrane potentials hyperpolarize approximately 0.5s to 1.5 s prior to the onset of the first swim cycle (Fig. 4.7a). In addition, depolarization of cell SIN1 during swimming generally terminates the swim episode. Although suppression of cell SIN1's activity is necessary for swimming, it is not sufficient to initiate swimming since hyperpolarization of cells SIN1 alone does not initiate swimming. A concurrent requirement is the activation of the segmental swim-generating network. Before swimming starts, spiking activity ceases in cell SIN1 while increasing in cell 204 (Fig. 4.7b).
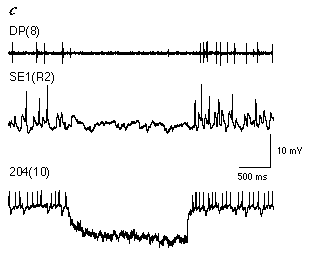
To date, only a few putative members of the swim-activating and inactivating systems have been identified (Brodfuehrer et al., 1995a). A potential candidate for the swim-activating system are cells SE1, a pair of interneurons in the subesophageal ganglion (Brodfuehrer et al., 1995b). Cells SE1 are generally spontaneously active, and receive feedback from the oscillator network during swimming. Their inclusion in the swim-activating system is based on the following observations: 1) Cell 204 receives direct excitatory input from cell SE1. 2) The level of excitation in cell 204 is positively correlated with the firing frequency of cell SE1. In fact, spiking activity in cell SE1 regulates the spontaneous level of activity in cell 204 to such an extent that when the swim motor program is not active, elimination of spiking activity in cell SE1 abolishes almost all EPSPs in cell 204 (Fig. 4.7c). 3) The level of excitation in both cell 115 and DE-3 is positively correlated with the activity of cell SE1. 4) Cell SE1 directly excites cells 28, 208, and DE-5. Thus cells SE1 have a profound excitatory influence on the swim-generating network.
On the otherhand, no strong candidtes for the swim-inactivating system have been identified. Cell SIN1 is most likely only a minor component of the swim-inactivating system since its activity level is controlled by other yet identified neurons, some of which are undoubtedly associated with leech behaviors that are incompatible with swimming such as whole-body shortening. Members of the swim-activating and -inactivating systems may also be part of a dynamic network that defines the 'internal state' of the leech nervous system, which is itself variable and modifiable, and influences the behavioral responsiveness of a leech to constant, repetitive stimuli (Grobstein, 1994).
Two observations document the existence of intrinsic variability within the leech nervous system, and that it affects the ability of a given input to elicit swimming. First, the motor output of isolated nerve cords intact from the head ganglion to the tail ganglion (H-T preparation) varies continuously in the absence of variations in input, but occassionally produces swimming without external input (Fig. 4.8a). Second, in an H-T preparation identical peripheral (dorsal posterior, DP) nerve stimulation sometimes, but not always, triggers swimming (Fig. 4.8b). There is no threshold stimulus voltage that consistently elicits swimming (Fig. 4.8c). In constrast, in a preparation consisting of a chain of ganglia from segmental ganglion 3 to the tail ganglion there is a clear threshold stimulus voltage where DP nerve stimulation reliably elicits swimming (Fig. 4.8c). Thus a property 'intrinsic' to the nervous system affects the behavioral responsiveness of leech preparations and is modifiable by changing the number of ganglia comprising the preparation; specifically by eliminating the influence of the head ganglion on the rest of the nervous system. This observation suggests that the ability of cell Tr1 stimulation to trigger swimming may depend upon the internal state of the nervous system.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}